


- 收藏
- 加入书签

香猪鼻腔细菌群落分析
 打开文本图片集
打开文本图片集
[ 通讯作者/第一作者E-mail:chenjiang654@gmc.edu.cn;]
【摘要】上呼吸道的鼻腔黏膜表面和其它与外界相通的黏膜组织一样,定植结构相对稳定和组成十分复杂的微生物菌群。鼻腔由于受到组织结构、生理功能以及外界诸多因素的影响,构成有别于呼吸道其他部位的独特生态环境。本研究克服传统培养法的缺陷,采用PCR-DGGE和RT-qPCR的方法,利用图象分析软件对DGGE图谱进行相关性分析。结果表明DNA酶结合CTAB法提取DNA更能客观反映鼻腔黏膜上定植的微生物多样性;处于微生物定植高峰期的二月龄香猪鼻腔黏膜内,细菌群落多样性指数明显高于其它月龄;稳定期的五月龄香猪,鼻腔细菌群落多样性指数最低;DGGE图谱中检测到大量未培养细菌,同时也检测到肺炎链球菌、副猪嗜血杆菌属、金黄色酿脓葡萄球菌属、流感嗜血杆菌属等常见的条件致病性微生物;感染组中香猪鼻腔内,金黄色葡萄球菌的拷贝数所占比例明显增加,可以作为鼻腔内呼吸道感染的致病性指示细菌。本文对香猪鼻腔中细菌群落结构的多样性、动态变化和稳定性进行分析,筛选出生境中导致香猪呼吸道感染的致病性指示微生物,为香猪呼吸道疾病的诊断和预防提供一定的理论参考。
【关键词】:微生物群落,PCR-DGGE,鼻腔,荧光定量PCR,香猪
1.背景
呼吸道疾病是导致香猪死亡的主要原因之一,上呼吸道的鼻腔黏膜系统上定植着许多共生菌、条件致病菌、致病菌,它们之间通过协同作用和拮抗作用,共同形成一个稳定的微环境1。各个黏膜组织作为一个“共同黏膜免疫系统”存在于呼吸道、口腔、泌尿生殖道、胃肠道。最新研究表明黏膜免疫系统作为一个统一的整体,某一局部黏膜系统内微生物群落失调,导致某些致病菌或者条件致病菌转化为优势菌属,致使疾病的产生2-4。目前猪的呼吸道疾病的诊断方法主要通过病原学检测,临床症状分析和免疫学检测。研究中发现这些方法并不能快速准确的给出诊断结果5-7。另外对未知病原菌无法进行检测,而且不能同时检测多种病源菌动态变化的情况8。
变性梯度凝胶电泳(DGGE )技术8用于检测DNA突变的电泳技术。基本原理是将具有化学变性剂梯度的聚丙烯酰胺凝胶用于区别的PCR扩增产物,产物长度相同而碱基排列不同的DNA片段会在相应的变性剂浓度下发生空间构型的差异,条带会停留在各自相应的变性剂梯度位置,形成DNA指纹图谱,获得物种之间DNA序列条带差异的一个轮廓,每条带代表一个特定的DNA序列,条带的数目和丰度呈现的是群落结构的多样性和丰富度。通过指纹图谱直观的观察到群落的多样性及动态变化,并对条带进行克隆序列进行种属鉴定。所以本实验采用变性梯度凝胶电泳和荧光定量PCR技术快速检测感染样本中所有疑似的病源菌,并且直观的判断其的动态变化,同时筛选出优势指示性病源菌比例,从而对呼吸道疾病及时诊断和治疗,并且为准确的治疗方案提供一定的理论依据。
2. 材料与方法
2.1主要材料和仪器
2.1.1 实验试剂
荧光定量PCR试剂(宝生物),pGEM-T Easy Vector Systems(普洛麦格),琼脂糖凝胶回收试剂盒(天根生化),质粒提取试剂盒(天根生化),质粒连接试剂盒(天根生化),引物合成(生工生物工程(上海)股份有限公司),SYBR Green I 染料(赛默飞),Goldview 染料(赛百盛)。
2.1.2 主要仪器
荧光定量PCR仪(Bio-Rad,CFX96 Touch),变形梯度凝胶电泳仪(Cleaver,VS20WAVE-DGGE),琼脂糖凝胶电泳仪(六一,DYCP-31DN),凝胶成像紫外透色系统(Bio-Rad,ChemiDoc XRS+)。
2.2 方法
2.2.1基因组提取:取鼻拭子样品用PBS 缓冲液悬浮刷洗,漩涡振荡,离心后收集细胞沉淀。使用CTAB/Nacl法提取样品中的DNA,加入TE缓存液重悬DNA,琼脂糖凝胶电泳检测提取的基因组质量及片段长度,核酸测定仪测定 DNA 浓度。
2.2.2 PCR 扩增反应及优化:用 341f和534r引物对扩增 16Sr RNA 基因 V3 区;用细菌通用引物27f 和1391r8扩增 16S r RNA 全长片段。利用Reconditioning PCR消除 PCR 过程产生的异源双链9;pGEM-T Easy Vector 通用引物扩增目的序列插入载体的目的片段。
2.2.3 DGGE 电泳:以16Sr RNA 基因 V3 区PCR扩增产物进行变性梯度凝胶电泳。凝胶成像仪成像,采用Quantity One 分析图谱上条带的数量、电泳位置和光密度值,通过UPGMA 算法进行聚类分析。
2.2.4 PCR 验证阳性克隆:DGGE 条带的切胶回收,DNA 片段的纯化和回收,连接反应,转化DH5α感受态细胞,阳性克隆子测序进行DGGE 图谱分析。
3 结果
3.1 样品的DNA提取及PCR优化
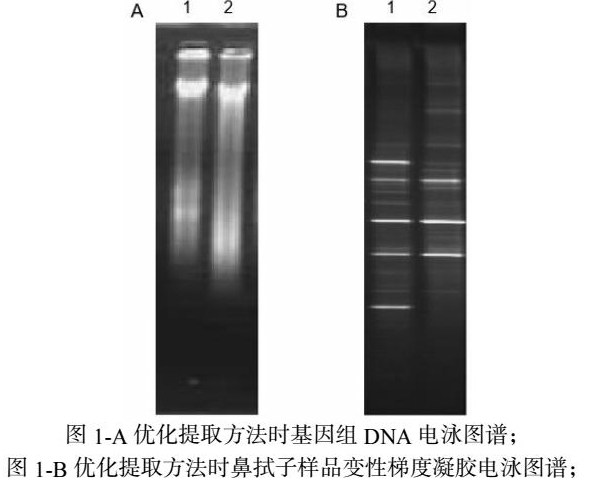
将提取的DNA样品进行琼脂糖凝胶和DGGE电泳检测。改进后的CTAB法10更能客观的反映样品中微生物群落。如图1-A 两种方法所获得的条带大小均匀一致,较常规CTAB法(条带2)提取获得的DNA,改进的CTAB法(条带1)获得的DNA条带较暗,但是DNA拖尾现象降低。图1-B经DGGE电泳两种方法均能获得条带丰富的图谱,样品1经DNA酶处理获得条带较少,说明改进后的CTAB法更能客观的反映样品中微生物群落。
3.2 16S rDNA V3区PCR-DGGE指纹图谱分析
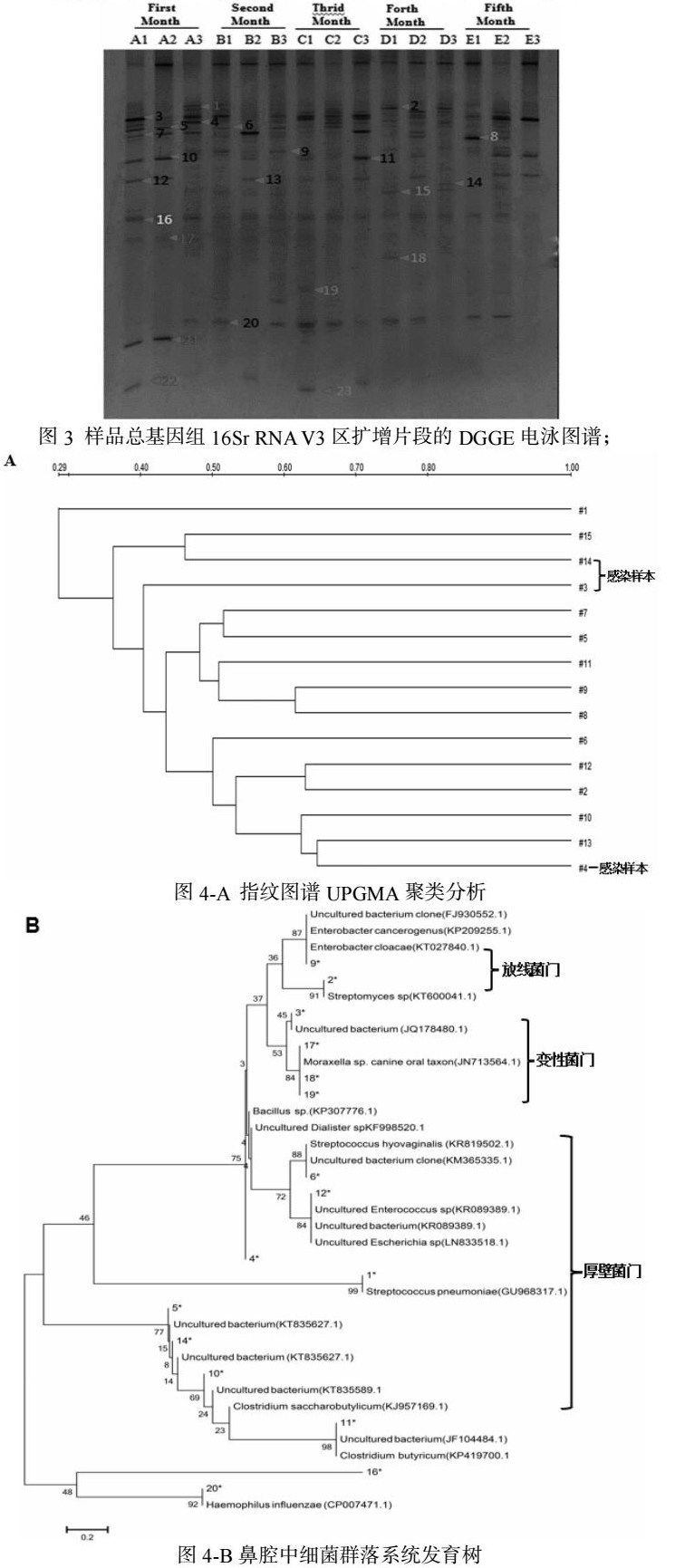
以提取的DNA为模板进行PCR扩增,第一轮常规PCR 以341f/534r为引物进行扩增,第二轮采用Touch Down-PCR以341f/534r 带GC夹的引物进行扩增,用2%琼脂糖凝胶电泳检测扩增结果如图2,两轮PCR均获得16S rDNA V3区特异性片段,产物长度180 bp左右,且条带亮度较高,可以满足后续 DGGE分析要求。利用变性梯度凝胶电泳(DGGE)分离16S rDNA V3区的特定长度的PCR产物15,不同样本的电泳图谱(图3)。图中各泳道中优势条带的电泳迁移位置一致。每个泳道中都同时存在多条亮度较高的 DNA 条带。通过DGGE图谱结合软件分析显示不同样品之间细菌群落扩增结果存在丰富的电泳条带且有一定差异;
3.3细菌DGGE图谱聚类分析
通过软件进行处理,并使用UPGMA对DGGE指纹图谱作聚类分析14(图4-A)。聚类分析细菌群落相似性结果:3、4、14感染样品DGGE图谱不存在明显的相似性。一月龄样品1、2和3之间群落相似性较低;二月龄样品4、5和6之间群落相似性较低;三月龄样品8和样品9群落相似性较高;四月龄样品10、11和12之间群落相似性较低;五月龄样品14和样品15群落相似性较高。对测序细菌构建系统发育树(图4-B),共克隆测序121个最终筛选出21个差异性结果,只有16.5%为已知菌,其余的均为未培养细菌占83.5%。所有的菌属被分别归属3个细菌门:厚壁菌门(Firmicutes)、变性菌门(Proteobacteria)、放线菌门(Actinobacteria)。
3.4优势病原细菌定量分析
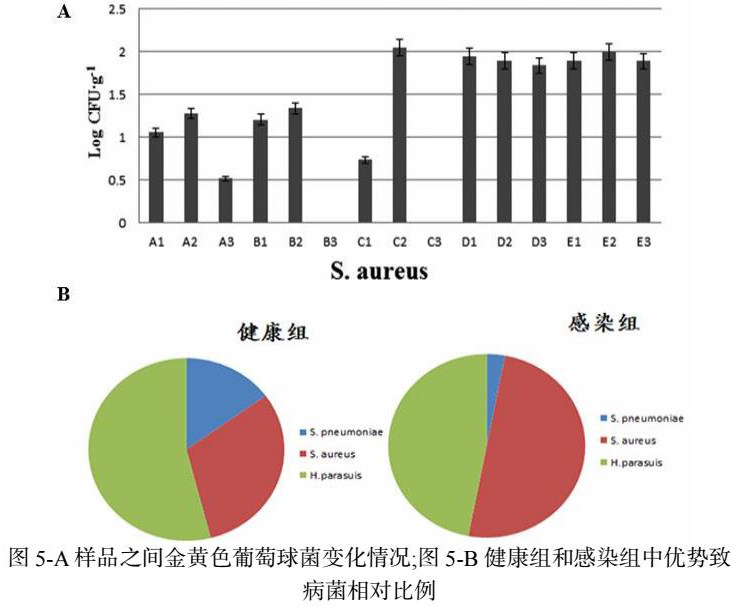
Blast比对发现S. pneumoniae 的 ply 基因,H. parasuis的ompP6 基因,S. aureus 的groL的基因分别为三种菌属特有基因11-13。针对这些基因,设计qPCR 引物。RT-qPCR对香猪鼻腔中金黄色葡萄球菌的定量分析:不同生长阶段的香猪鼻腔中金黄色葡萄球菌定量检测,结果变化趋势(图5-A),金黄色葡萄球菌拷贝数变化在0-2.04之间,平均值为1.303±0.71,各个组间金黄色葡萄球菌含量有差别,其中C2含量最高,B3、C3含量最低。感染组之间,拷贝数变化在0-1.99之间,平均值为1.233±0.74。健康组之间,拷贝数变化在0-2.04,平均值为1.217±0.77,进行统计检验,发现差异不显著P=0.858(P>0.05)。但是感染组中S. aureus 的拷贝数比例相对增S.pneumoniae 和 H.parasuis 的拷贝数比例相对减少(图5-B)。
4. 讨论
本研究通过改进的CTAB法提取香猪鼻腔微生物菌群基因组DNA,利用通用引物进行扩增,产物经DGGE电泳,通过相关性分析软件对其群落DGGE图谱中条带相似性系数和多样性指数进行分析。结果表明:香猪鼻腔中肺炎链球菌、金黄色酿脓葡萄球菌属、莫拉克斯氏菌属、流感嗜血杆菌属、副猪嗜血杆菌属均属于呼吸道常见条件致病性细菌;其中鼻腔中肺炎链球菌、金黄色酿脓葡萄球菌属和副猪嗜血杆菌属三种条件致病性细菌优势菌属都存在于健康组与感染组、不同生长阶段的香猪鼻腔中数量差异不显著(P>0.05);处于微生物定植时期的二月龄香猪,鼻腔细菌群落多样性指数明显高于其它月龄;生长稳定期的五月龄香猪,鼻腔细菌群落多样性指数最低;感染组中香猪鼻腔中金黄色葡萄球菌数量相对比例明显增加;本研究中香猪呼吸道感染的指示性病原菌是金黄色葡萄球菌,可针对其引起的感染提出合理的治疗方案。
参考文献
[1] Alvarez A H, Gutiérrez-Ortega A, Hernández-Gutiérrez R. Identification of immunodominant proteins from Mannheimia haemolytica and Histophilus somni by an immunoproteomic approach[J]. Canadian Journal of Veterinary Research, 2015, 79(4): 339-342.
[2] Amann R I, Ludwig W, Schleifer K H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation.[J]. Microbiological Reviews, 1995, 59(59): 143-169.
[3] Beck-Sagué C M, Jarvis W R. Secular trends in the epidemiology of nosocomial fungal infections in the United States, 1980-1990[J]. The Journal of infectious diseases, 1993: 1247-1251.
[4] Porebski S, Bailey L G, Baum B R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components[J]. Plant molecular biology reporter, 1997, 15(1): 8-15.
[5] Qiu X, Wu L, Huang H, et al. Evaluation of PCR-generated chimeras, mutations, and heteroduplexes with 16S rRNA gene-based cloning.[J]. Applied & Environmental Microbiology, 2001, 67(2): 880-887.
[6] 曹鹏,贺纪正. 微生物生态学理论框架[J]. 生态学报,2015,35(22):7263-7273.
[7] 高启禹,徐光翠,李小英. 变性梯度凝胶电泳 (DGGE) 在微生物多样性中的研究[J]. 生物学杂志,2009,26(5):80-82.
[8] 韩莹,刘宏伟,刘洋等. 念珠菌在艾滋病患者龈沟液中的检出率[C],2008.
[9] 冉玉平,刘素玲. 利用核糖体RNA(rDNA)基因序列分析法鉴定真菌菌种[C],2007.
[10] 束弋,王健伟. 肠道黏膜免疫系统的研究进展[J]. 微生物与感染,2007,2(1):59-62.
[11] 王剑,李凤琦等. 呼吸道共生菌诱导M2型肺泡巨噬细胞减轻流感病毒感染导致的免疫损伤[C],2012.
[12] 钟锋. 下呼吸道感染金黄色葡萄球菌耐药性监测[D]:安徽医科大学,2012.
[13] 邹莎莎,程安春,汪铭书. 肺炎链球菌毒力因子的研究进展[J]. 中国人兽共患病学报,2009,25(4):372-375.
[14] 叶涛. 物种多样性综合指数研究[D]:中南林业科技大学,2014
[15] Zhang X, Yan X, Gao P, et al. Optimized sequence retrieval from single bands of temperature gradient gel electrophoresis profiles of the amplified 16S rDNA fragments from an activated sludge system[J]. Journal of Microbiological Methods, 2005, 60(1): 1-11.

 京公网安备 11011302003690号
京公网安备 11011302003690号


