


- 收藏
- 加入书签

杉木植株几种生理生化性状与其种子质量产量的关系探讨
 打开文本图片集
打开文本图片集
摘 要 本文通过测定杉木一代种子园45个无性系植物的蒸腾强度及枝叶中多酚氧化酶、单宁、铁、硼、铜的含量,同时与无性系相应的单株产量、种子发芽率、简化活力、出籽率建立回归关系,进行相关显著性检验,结果表明:铁、铜、硼元素的含量与种子质量产量关系显著,铁、硼对提高种子的产量有显著效果。铜降低单株产量,但可提高出籽率。单宁、多酚氧化酶发芽率呈显著的负相关系,蒸腾强度在一定范围内提高有利于提高单株产量,通过改变微量元素含量,改变代谢方向,达到提高杉木种子质量产量的目的。
关键词 杉木 生理生化 种子质量产量
杉木是我国南方重要的速生乡土用材树种,人工林面积和蓄积量分别占全国人工乔木林的1/5和1/4,造林面积大,种苗需求量多,但在杉木种子中,健全粒种子只有30~50%,其他是涩粒、空粒或腐烂种子等异常种子[1,2],严重影响了杉木种子的质量与产量,而杉木种子的质量与产量同植株发生的生物化学过程有关。通过对45个不同无性系植物的蒸腾强度及枝叶中多酚氧化酶、单宁、铁、硼、铜的含量进行分析,从生理生化角度来寻求提高杉木种子的质量和产量;为其提供理论依据,并从生理生化角度着手,力求从调节母树养分状况,改变代谢方向,降低种子败育率,提高杉木种子园的质量与产量。
1 材料与方法
1.1 材料
1.1.1 自然条件 试验地设在福建省尤溪县国有林场华口溪良种基地一代种子园,位于东经118°12′,北纬26°15,海拔150-200m,坡度20°-30°,年平均气温18.9° C,年降水量1579mm,无霜期299-322d,属戴云山脉山地地区,土壤为红壤,土层80-100cm,表土层5-10cm,土壤肥力中等。1985年建立一代种子园,1987年嫁接,1991年开始生产种子,密度为300-450株/hm2,郁闭度0.5-0.6,收集了45个优良无性系,采用集约经营管理。
1.1.2 材料选择 从该良种基地45个无性系中每个无性系选择1单株,三个重复,共135个单株,于四、五月份从树冠中间选择生长健壮正常的枝条进行生理生化分析。十一月中旬在各无性系单株中采集种子,分别单株处理。用解剖法进行养分含量分析。
1.2 方法
1.2.1 蒸腾强度的测定 利用蒸腾吸水量测定,根据氯化钴纸在干燥时为蓝色,当吸收水分后,变为粉红色,根据变色所需时间长短,然后按钴纸标准吸水量计算出作物蒸腾强度。
1.2.2 微量元素含量测定 供试样品烘干粉碎,进行各项测定[3]
1.2.2.1 铁 灼烧提取后,用磷酸—草酸一磺基水杨酸法测定,根据在不同酸度下Fe3和磺基水杨酸生成不同的稳定配合物,因溶液的PH不同,形成配合物的组成也不同,来测定铁的含量。
1.2.2.2 铜 对苯硫腙一四氯化碳溶液提取, 用二乙基二硫代甲酸钠比色法测铜含量,根据铜与二乙基二硫代氨基甲酸钠在氢氧化铵溶液中生成黄棕色胶体络合物,此络合物易溶于有机溶剂,用四氯化碳萃取比色测定铜含量。
1.2.2.3 硼 灼烧提取后,用洋红一硫酸比色法测定,系在浓硫酸中使硼和有机试剂络合,生成有色物质,再用有机溶剂萃取有色络合物,进行分光光度测定硼含量。
1.2.3 单宁含量 用磷钼酸一钨酸钠比色法测定[4]。用丙酮溶液提取作物中单宁,溶液经过滤后,取滤液加钨酸钠-磷钼酸混合溶液,显色后,在760 nm波长处测定吸光度值,标准曲线法测定作物中单宁含量。
1.2.4 多酚氧化酶活性测定 用氧化邻苯二酸一抗坏血酸碘酸钾滴定法测定[3],利用邻苯二酸在多酚氧化酶催化下受氧作用生成邻苯二醌,邻苯二醌能够被抗坏血酸氧化还原,把分析对象配成PH6左右的样液,用碘酸钾进行滴定,测得剩余的抗坏血酸,由得到的数据求出被氧化的抗坏血酸量,并计算出多酚氧化酶活性。
1.2.5 发芽率的测定
种子发芽实验按GB2772-1999《林木种子检验方法》进行, 种子发芽=种子发芽数/供试种子数*100%,计算发芽率,发芽试验结束后,将幼根切下,放在98℃-102℃烘至恒重,测定各供试样品的幼根干重,计算简化活力。简化活力指数G·S值=发芽率(量)×生长势(根重)[5]。
1.2.6 种子产量测定
各无性系单株产量于l1月中旬分单株采种称重测定产量,各无性系单株从树冠中部及东西南北各取4个球果装入信封中,阴干后取出全部种子,用解剖法测定种子的质量,解剖时按按优良种子、涩粒、空粒、腐烂粒分别记录,计算出籽率。
1.2.7 分析统计方法
测得多酚氧化酶,单宁,铁、铜、硼等微量元素与单株产量、发芽率、简化活力、出籽率进行回归分析,计算相关指数,并以95%可靠性进行显著性检验,测得多酚氧化酶、单宁和铁、铜、硼等微量元素与产量、发芽率、简化活力、出籽率之间的关系。
2 结果与分析
2.1 微量元素与杉木种子质量产量的相关分析
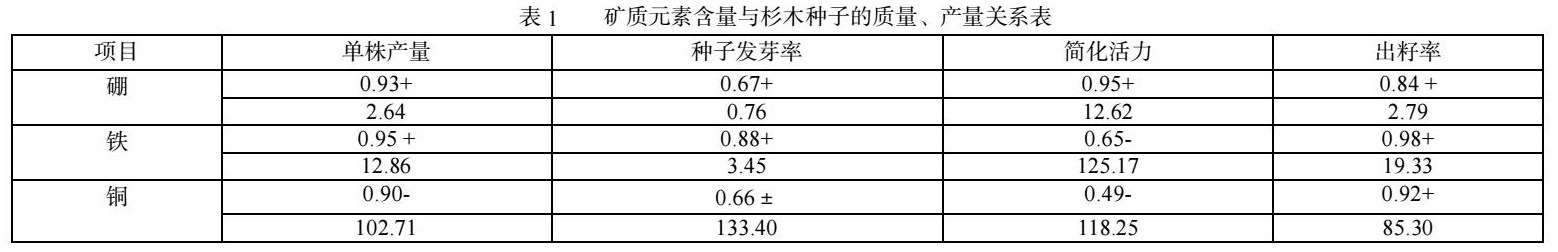
植物从外界吸收营养,不同的土壤环境下的植物其成分组成,含量不同,即使是在同样的土壤条件下,不同的植株由于营养元素的富集力不同,其成分组成,含量也各不相同,植物体内的矿质元素,一部分用来建造自身的结构物质,另一部分用来参与酶促反应,能量代谢和生理调节,因此各元素含量是影响杉木种子形成的重要因素,根据对各无性系有关微量元素进行测定统计分析,结果见表1。
硼在植物体内的功能主要是加快酶代谢和木质素形成,参与碳水化合物运输与蛋白质代谢,促进花粉的萌发和花粉管的伸长,减少花而不实和花粉畸形现象,避免花、蕾易脱落,受精不正常,果实种子不充实等。花粉母细胞不能参与四分体的分化。因此,由于硼能显著地促进开花结实、传粉、授精,固硼能提高种子的单株产量,同时,由于硼可以减少种子早期败育,降低种子的空粒率,因此使出籽率得到提高,相关分析的结果证实了这一点。硼会影响酚的合成,缺乏硼会使呼吸代谢转向戊糖支路,为酚提供更多的底物,适量的硼能正常结合过量的酚酸,避免酚酸造成的毒害。缺硼必然导致酚含量的提高,因而适当提高硼含量,有利于提高种子单株产量和出籽率。
铁是植物光合作用、生物固氮、和呼吸作用中的细胞色素和非血红素铁蛋白的组成,是植物有氧呼吸的酶的重要组成物质,是植物能量代谢的重要物质,缺铁影响植物生理活性,也影响养分吸收。铁与杉木种子的质量产量形成密切相关,表中铁的含量与单位株产量、种子发芽率、出籽率存在着显著的正相关,铁对合成叶绿素是必需的,因而铁的含量高时能提高单产量,在生理功能上,铁能作为一些酶如过氧化物酶,细胞色素氧化酶的组成成分,因此含铁高的植物,其单宁合成也比较旺盛,含量过高会促进涩粒产生。
铜的主要作用是作为许多种氧化酶及质体蓝素的构成部分,对调节呼吸作用和光合作用有重要作用,它能促进蔗糖等碳水化合物向生殖器官流动,促进植物的生长发育,有利于花粉发芽和刺激花粉管伸长,适当增加铜,可以促进花粉受精,有利于优良种子形成。在缺铜的情况下,常因生殖器官的发育受到阻碍,而使植株发生某些生理病害,引起植物发育不全,甚至不能结籽,空籽粒多。但植物体对铜的需要量较低,过量的铜易造成毒害,相关分析表明,铜的含量与单株产量成负相关,与种子的出籽率呈正相关。
2.2 单宁、多酚氧化酶、蒸腾强度与产量、质量的相关分析
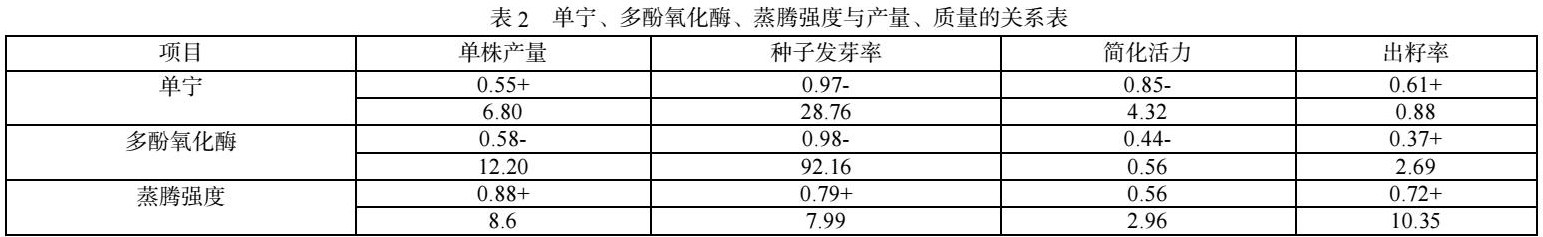
从各个无性系中测得枝条的蒸腾强度、单宁、多酚氧化酶的含量,与相应的单株产量发芽率、简化活力、出籽率进行回归分析,以95%的可靠性进行相关显著性检验、根据曲线变化趋势,确定相关的正负性,结果见表2
从表2可以看出,杉木植株中单宁与杉木种子的发芽率,呈显著的负相关,从单宁对植物本身意义来说,单宁不仅仅是一种代谢废物,它在一定程度上可以抑制病毒、微生物、病虫害的活动,因此对球果的形成、降低早期败育有一定作用,但是单宁的积累最终必然导致内部生理平衡的破坏,由于对涩粒的形成影响很大,所以与种子的发芽率形成显著的负相关。
相关分析表明,杉木植物中多酚氧化酶的含量与种子的发芽率呈负相关,一般认为,多酚氧化酶直接参与单宁的生成,可以氧化多元酚为醌,在多酚氧化酶作用下,植物体内黄烷-3-醇类物质脱氢氧化为邻醌,然后发生头与尾型,或尾与尾型的结合,这种反应不断地继续,产生二聚体、三聚体、多聚体。产物由水解类单宁到凝缩类单宁。单宁特别是凝缩类单宁含量的积累,在酸作用下产生红色沉淀,破坏了细胞生理代谢导致涩粒形成,直接影响了种子的质量与产量。此外,多酚氧化酶的活动使种子中大量养分转化为单宁,不利优良种子的发育,一定程度上导致了单株产量下降。
对蒸腾强度进行分析的结果表明,蒸腾强度在一定范围内提高有利于提高单株产量。由于对种子优良率、涩粒、空粒的影响,也间接影响了种子的发芽率、简化活力。蒸腾强度大,植物代谢旺盛,养分得以输送,这对提高杉木种子的产量与质量有一定作用。
3 小结
3.1 铁、铜、硼元素的含量与杉木种子单位株产量、种子发芽率、简化活力、出籽率关系显著,硼与这几项指标呈正相关关系,硼对提高种子产量,减少涩粒有归显著的效果;铁的含量与单位株产量、种子发芽率、出籽率存在着显著的正相关,可以提高种子发芽率、出籽率;铜的含量与单株产量成负相关,与种子的出籽率呈正相关,可以减少种子涩粒的形成。
3.2 单宁、多酚氧化酶与杉木种子的发芽率,呈显著的负相关,单宁的积累是导致涩粒形成的重要原因,抑制多酚氧化酶的活性,可以一定程度上阻碍单宁的生成。降低种子的涩粒率,提高种子的质量与产量。
3.3 植物体内生理活动是一个综合的整体,各种因素并非单独存在,而是相互作用,互相调节,生产上我们应该综合分析各种因素,因地制宜,选择优良的无性系,合理地施用含硼、铜元素的肥料,少施含铁肥粒,使母树的营养生长与生殖生长协调一致。由于微量元素对植物的生命活动影响非常巨大,而土壤往往又不能完全及时满足作物的需要。科学施肥可以改变作物代谢功能,促进作物体内蛋白质、淀粉、糖分、脂肪、生物碱和其他物质积累,因而,科学施肥就成为提高种子产量和改良品质的主要措施之一。
参考文献
[1]叶培忠.陈岳武.蒋恕等.杉木种子生活力变异的研究[J].南京林产工业学院学报,1981,(3):22~32;
[2]管康林. 杉木种子空瘪涩粒成因的综合分析 [J].浙江林学院学报,1997,14(1):88~93;
[3] (美)J 帮纳,J E 瓦纳编;植物生物化学翻译组译.植物生物化学[M].北京;科学出版社,1984:1~4,48~52,55~61,197~209;
[4]中国土壤学会农化专业委员会编.土壤农业常规化学分析方法[M].北京:科学出版社,1983: 343~345;
[5]顾增辉.徐本美.郑光华. 测定种子活力方法的掠讨(2)— 发芽生理涮定法[J].种子,1982,(3):11~17;
[6]许鲁平. 第3代杉木种子园产量与土壤微生物关系的研究[J] .南昌工程学院学报. 2013,(4):38~41;
[7]王赵民.张建忠.倪荣新等 . 杉木种子园产量和品质的影响因子分析 [J]. 浙江农林大学学报,1998,(1):13~21;
[8]翁春媚.齐明.华朝晖等. 杉木播种品质的遗传分析与空瘪涩粒成因探讨[J].福建林业科技,2016(2):160~165;
[8]胡德活.阮梓材.陈仲.杉木种子园种子产量及其相关因子的分析[J].林业与环境科学,1994(3):42~46;[9]刘文飞.吴建平.孟庆银.等 杉木第3代种子园母树和土壤营养元素分析[J]. 福建林业科技,2013(1):70~74;
[10]高智慧.史忠礼.许月明等.杉木种子发育生理、涩籽成因及降低败育措施的研究[J]. 林业科学研究,2000,13(6):659~666

 京公网安备 11011302003690号
京公网安备 11011302003690号


