


- 收藏
- 加入书签

iRhom2在多种疾病中的作用机制及研究进展
 打开文本图片集
打开文本图片集
摘要 非活性菱形蛋白2(Inactive rhomboid protein 2,iRhom2)由Rhbdf2基因编码,虽没有蛋白酶活性,但是参与调控细胞内蛋白如肿瘤坏死因子α转化酶( Tumor necrosis factor-a converting enzyme,TACE)即ADAM17、干扰素基因刺激蛋白(Stimulator of interferon genes,STNG)等从内质网到高尔基体、核周微粒体的运输与活化过程,在炎症相关疾病和病毒介导的固有免疫中发挥重要作用。此外iRhom2在出血性疾病、代谢性疾病和关节病中亦具有关键影响。本文将对iRhom2在这些疾病中扮演的角色及相关作用机制进行综述。
关键词 非活性菱形蛋白2;炎症反应;肿瘤坏死因子α转换酶(ADAM17); 新型多功能蛋白
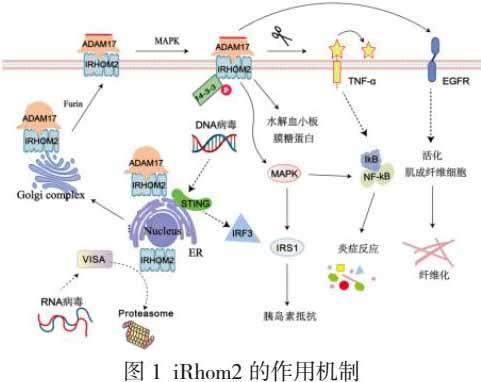
非活性菱形蛋白2(Inactive rhomboid protein 2,iRhom2)是由Rhbdf2基因编码的。iRhoms作为Rhomboid家族无蛋白酶活性的一类成员,具有6个iRhoms同源结构域、关键的催化域和细胞内的N端结构域,其N端结构域(aal-403)是iRhom2与其他蛋白相互结合的关键区域。其中iRhom1在全身多处组织表达,而iRhom2则主要在免疫系统细胞和上皮细胞等部位表达[1],其主要分布在内质网。作为一种新型多功能蛋白,对细胞生长和机体功能发挥积极作用,然而当Rhbdf2基因发生突变或蛋白异常表达时,也可能产生病理性影响[2]。
最初科学家发现哺乳动物体内的iRhom2蛋白通过耦合并运输肿瘤坏死因子α转化酶(TACE)即ADAM17对炎症因子的释放至关重要,可调节凋亡和炎症反应。在该过程中,iRhom2蛋白结合并转运内质网中的ADAM17至高尔基体,然后ADAM17被活化并释放到质膜[3-4]。最终,活化的ADAM17切割活化许多生物分子,如肿瘤坏死因子α (TNF-α)、表皮生长因子受体(EGFR)、白细胞介素6 (IL-6) 等[5]。最近研究报道iRhom2的失活阻止免疫细胞成熟,而不影响组织中的稳态[6],因此iRhom2成为了治疗TNF-α相关疾病的理想靶点。
1.iRhom2与非酒精性脂肪性肝炎
非酒精性脂肪性肝病(NAFLD)是指在排除酗酒和药物损伤等明确损肝因素的情况下,有超过5%肝细胞中出现脂质过度积累的临床病理综合征。非酒精性脂肪性肝炎(NASH)作为NAFLD中较严重的亚型。其典型病理特征是:肝细胞呈现脂肪变性和气球样改变,伴有炎症,可能进一步发展为纤维化[7],但是目前仍没有针对该疾病的特效上市药[8]。
iRhom2蛋白不仅能够通过耦合并运输ADAM17影响炎症细胞因子,达到促进炎症的作用。在NASH的发展过程中还有多种更加复杂的功能。谭君教授[9]等首次证实,高脂饮食(HFD)长期摄入诱发NAFLD和NASH病理表型的主要起始分子靶点是iRhom2。一方面其直接与MAP3K7相互作用,上调了MAP3K7–NF-κB途径,在肝细胞和Kupffer细胞中发挥促炎作用。其次iRhom2能够参与肝细胞糖脂代谢过程,促进JNK/IRS1信号的激活,但干扰AKT/GSK3β相关的胰岛素信号[10],在肝实质细胞中发挥着重要作用。紧接着课题组通过体外转染实验表明Trim31通过K48连接的多聚泛素化能够促进iRhom2的降解,使小鼠血清中的炎性介质水平显著降低,从而减轻肝脂肪变性、炎症和肝纤维化[11]。相反,去泛素化酶USP13能够移除iRhom2的K63泛素化修饰,从而降低该蛋白稳定性和胞内丰度,因此抑制iRhom2及其下游信号通路的活化水平,进一步抑制NASH病理表型的发展[12]。由此可见,iRhom2蛋白丰度与NASH病损程度密切相关。
2.iRhom2与出血性关节病
出血性血小板病患者的iRhom2和ADAM17蛋白表达水平明显高于健康者[13]。出血性血小板病发病过程中,iRhom2通过与pro-ADAM17结合,激活该信号通路,从而介导血小板膜表面的血小板膜糖蛋白(GP)Ib / IX /V胞外功能区过度水解,引起血小板凝血酶受体复合物活性下降,导致血小板聚集缺陷,皮肤黏膜出血。益气活血方和消淤止血片抑制出血性血小板病患者ADAM17对血小板膜表面蛋白水解功能,可能是通过调控其上游iRhom2表达、功能或者活性而实现的[14-15]。
血友病是由于凝血因子VIII/IX缺乏症引起的一种先天性出血性疾病,呈X染色体连锁。其关节出血后的关节病变仍未得到有效防治,影响着患者的身心健康。充血对关节软骨具有直接侵蚀作用。一方面,关节腔充血后,激活单核/巨噬细胞释放TNF-α、基质金属蛋白酶类等细胞因子,导致血管内皮生长因子(VEGF)分泌增加,诱发滑膜肥大和新生血管生长,形成滑膜炎症。此外,出血后造成铁沉积,诱导关节软骨细胞损伤和死亡,最终造成整个关节的功能障碍[16]。在K/BxN模型小鼠中删除骨髓细胞ADAM17能减少类风湿关节炎的发生,而缺乏iRhom2也能预防关节病形成[17]。在血友病关节病进展过程中,血液进入关节,iRhom2即可促进TNF-α释放。原发性骨质疏松症伴骨痛患者的TNF-α也明显高于正常者,与iRhom2/TACE 信号通路异常表达相关[18-19]。
3.iRhom2与肾损伤肾纤维化
急性肾损害(AKI)是由于肾脏功能在较短时间内突然降低而导致的临床综合症。主要表现为肌酐、尿素氮等含氮代谢产物在血液中显著增加,水钠潴留、电解质失调、酸碱失衡等,以较高的发病率和病死率为特征,易发展为慢性肾脏病。与正常小鼠相比,iRhom2敲除可保护PM2.5暴露的小鼠免于发展为严重的肾损伤[20],并改善了肾脏的病理变化。这可能至少部分是通过抑制TNF-α/NF-κB信号通路,此外,还通过增强血红素氧合酶1/核因子-E2相关因子2 (HO-1/Nrf-2)的表达,降低c-Jun氨基末端激酶(JNK)激活,抑制氧化应激[21]。
发生急性肾脏损伤后成纤维化细胞就会发生迁移、增殖并最终分化为肌成纤维细胞(MFBs),这些细胞会参与到肾小管的纤维化疤痕形成中。EGFR的激活对于肌成纤维细胞的活化、迁移和增殖非常必要。而在细胞水平上,EGFR能被ADAM17所激活。因此iRhom2缺失的小鼠体内EGFR的激活较少,并且在肾损伤后较少发生肾纤维化[22]。
4.iRhom2与病毒感染
VISA(又称MAVS)是先天免疫应答RNA病毒的重要接头蛋白。在病毒感染前后,VISA蛋白水平受到精细调控,以确保先天抗病毒反应的最佳激活和及时终止。研究发现iRhom2通过不同的机制维持VISA的稳定性,在对RNA病毒进行有效的先天免疫应答中发挥重要作用。在未感染和早期感染时,iRhom2介导E3泛素连接酶RNF5的自泛素化和降解,损害VISA-RNF5-GP78复合物的组装,从而拮抗相关降解。病毒感染晚期,iRhom2介导E3泛素连接酶MARCH5的蛋白酶体依赖性降解,损害VISA的线粒体相关降解。iRhom2作为VISA的检查点,确保了对RNA病毒的适当先天免疫反应[23]。
舒红兵研究组[24]首次报道了iRhom2通过介导接头蛋白STING的转运和稳定性在抗DNA病毒天然免疫中发挥关键作用。实验表明,iRhom2通过两种途径调控STING。一方面,iRhom2通过桥接转运肿瘤坏死因子受体相关蛋白TRAPβ到STING,促进其从内质网转运到核外周区域的微体,并在此过程中激活下游的转录因子IRF3;另一方面,iRhom2招募去泛素化酶EIF3S5,通过去除STING的K48泛素化链,维持该蛋白水平的稳定性和丰度,促进STING对下游转录因子IRF3的激活[25]。
5.结论与展望
总之,iRhom2作为一种新型多功能蛋白,与多种疾病有关。早在2012年就首次报道了iRhom2对于ADAM17的成熟和运输至关重要[26],确定了其作为抗炎的重要靶标。之后研究发现,iRhom2通过在STING复合体上募集TRAPβ ,促进其从内质网转运到核周微粒体[27],在DNA病毒诱导的先天免疫中发挥重要作用。此外iRhom2蛋白通过多种信号通路成为治疗疾病的理想靶点,如NASH、出血性关节炎[28]、肾损伤及纤维化等(如图1)。iRhom2蛋白调节机制错综复杂,除上述所述外还涉及其他多个信号通路,目前研究所揭示的仅是冰山一角,不管是对炎症类疾病的共性调节机制还是不同疾病的个性调控都有待于进一步的研究。
参考文献
[1] LEE M Y, NAM K H, CHOI K C. IRhoms:its functions and essential roles [J]. Biomol Ther (Seoul), 2016,24(2):109-114.
[2] BERGBOLD N, LEMBERG M K. Emerging role of rhomboid family proteins in mammalian biology and disease [J]. Biochim Biophys Acta, 2013, 1828(12): 2840-2848.
[3] DÜSTERHÖFT S, BABENDREYER A, GIESE A A, et al. Status update on iRhom and ADAM17: it's still complicated [J]. Biochim Biophys Acta Mol Cell Res, 2019, 1866(10): 1567-1583.
[4] 徐榕, 康海锋, 刘宏斌,等. 去整合素-金属蛋白酶17在消化系统恶性肿瘤中的研究现状[J]. 世界华人消化杂志, 2020, 28(19): 986-991.
[5] AL-SALIHI M A, LANG P A. IRhom2: an emerging adaptor regulating immunity and disease [J]. Int J Mol Sci, 2020, 21(18): 6570-6590.
[6] LICHTENTHALER S F. IRHOM2 takes control of rheumatoid arthritis [J]. J Clin Invest, 2013, 123(2): 560-562.
[7] 杜重锦, 张晓慧, 孔烁,等. 从非酒精性脂肪性肝病到代谢相关脂肪性肝疾病 [J]. 安徽医药, 2024, 28(05): 1039-1043.
[8] 田长林, 李玉琴, 刘洪涛. 非酒精性脂肪性肝病新药的临床研究进展 [J]. 中国现代应用药学, 2024, 41(06): 858-863.
[9] 谭君, 徐敏轩, 葛晨旭,等. 非酒精性脂肪性肝病(NAFLD)治疗新靶点IRHOM2的发现与天然药物干预新机制 [J]. 中国科技成果, 2024, 25(3): 51-53.
[10] XU M, GE C, ZHU L, et al. IRhom2 promotes hepatic steatosis by activating MAP3K7-dependent pathway [J]. Hepatology, 2021, 73(4): 1346-1364.
[11] XU M, TAN J, DONG W, et al. The E3 ubiquitin-protein ligase trim31 alleviates non-alcoholic fatty liver disease by targeting Rhbdf2 in mouse hepatocytes [J]. Nat Commun, 2022, 13(1): 1052-1072.
[12] XU M, TAN J, ZHU L, et al. The deubiquitinating enzyme 13 retards non-alcoholic steatohepatitis via blocking inactive rhomboid protein 2-dependent pathway [J]. Acta Pharm Sin B, 2023, 13(3): 1071-1092.
[13] 杨胜兰, 沈霖, 张红星, 等. 脾虚型出血性血小板病患者ADAM17的表达变化及健脾益气活血方的治疗作用 [J].中国中西医结合消化杂志, 2015, 23(02): 75-78.
[14] 卢芙蓉, 秦铀, 沈霖,等. 基于iRhom2/ADAM17信号通路探讨益气活血方治疗出血性血小板病患者的作用机制 [J]. 中西医结合研究, 2020, 12(04): 217-221.
[15] 沈霖, 沈迪, 朱闽,等. 消瘀止血片对出血性血小板病患者凝血酶受体GPⅠb/Ⅸ/Ⅴ及GPⅠbα表达的影响 [J]. 中国中西医结合杂志, 2008, 28(12): 1086-1089.
[16] ASTERMARK J, DOLAN G, HILBERG T, et al. Managing haemophilia for life: 4th haemophilia global summit [J]. Haemophilia, 2014, 20(5): 1-20.
[17] 刘国青, 陈振萍, 吴润晖. 血友病关节病变与炎症因子关系的研究进展 [J]. 血栓与止血学, 2016, 22(6): 718-720.
[18] 徐晓娟, 沈霖, 杨艳萍. 密盖息对骨质疏松患者血清中IGF-1、IL-6以及TNF-a水平的影响[J]. 中国疼痛医学杂志, 2011, 17(11): 648-653.
[19] 帅波, 马陈, 沈霖,等. 原发性骨质疏松症伴中、重度骨痛患者iRhom2/TACE信号通路表达 [J]. 中华骨质疏松和骨矿盐疾病杂志, 2020, 13(06): 535-538.
[20] 白丽荣, 谭子康, 龚航远,等. 大气PM2.5全身暴露对大鼠多脏器病理, 氧化应激指标和炎症因子的影响 [J]. Asian Journals of Ecotoxicology, 2020, 15(6): 132-140.
[21] CHENXU G, MINXUAN X, YUTING Q, et al. IRhom2 loss alleviates renal injury in long-term PM2.5-exposed mice by suppression of inflammation and oxidative stress [J]. Redox Biol, 2018, 19:147-157.
[22] CAO S, PAN Y, TERKER A S, et al. Epidermal growth factor receptor activation is essential for kidney fibrosis development [J]. Nat Commun, 2023, 14(1): 7357-7374.
[23] LUO W-W, LI S, LI C, et al. IRhom2 is essential for innate immunity to RNA virus by antagonizing ER-and mitochondria-associated degradation of VISA [J]. PLoS pathogens, 2017, 13(11): e1006693.
[24] LUO W-W, LI S, LI C, et al. iRhom2 is essential for innate immunity to DNA viruses by mediating trafficking and stability of the adaptor STING [J]. Nature immunology, 2016, 17(9): 1057-1066.
[25] 张雪, 李曼, 张鑫,等. 干扰素基因刺激蛋白在慢性肝病发生发展中的抑制作用 [J]. 临床肝胆病杂志, 2019, 35(5): 1148-1152.
[26] 江海龙, 王宁远, 陆一鸣. 肿瘤坏死因子受体选择性拮抗剂的研究进展 [J]. 药学实践杂志, 2015, 33(5): 392-395.
[27] 徐赫男, 李新宇, 方敏, 等. TANK结合激酶1在固有免疫应答中的作用及其泛素化调控 [J]. 生物工程学报, 2021, 37(4): 1189-1204.
[28] 林振洋, 王冶凡, 张飞旭, 等. AAV介导的TNFR关节内基因治疗对血友病性关节病预防和治疗的实验研究 [J].华东理工大学学报 (自然科学版), 2022, 48(5): 631-640.
作者简介:刘鑫,女,硕士研究生 研究方向:药理学

 京公网安备 11011302003690号
京公网安备 11011302003690号


